
Tue, Jun 23, 2026
[Archive]
Volume 5, Issue 3 (Aug 2020)
JNFS 2020, 5(3): 248-258 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Fallah Tafty S, lak S, Mojaddam M, Naderi A, Abdollahian-Noghabi M. The Effect of Different Doses of Tank-mixed Herbicides on Antioxidant Enzymes Activity of Soybean. JNFS 2020; 5 (3) :248-258
URL: http://jnfs.ssu.ac.ir/article-1-288-en.html
URL: http://jnfs.ssu.ac.ir/article-1-288-en.html



Department of Agronomy, Ahvaz Branch, Islamic Azad University, Ahvaz, Iran
Full-Text [PDF 731 kb]
(1070 Downloads)
| Abstract (HTML) (3418 Views)
.png)
.png)
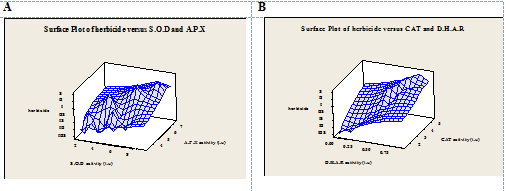
Figure 3. Effect of different herbicide doses on CAT and DHAR activity
Full-Text: (1003 Views)
The Effect of Different Doses of Tank-mixed Herbicides on Antioxidant Enzymes Activity of Soybean
Sina Fallah-Tafty; MSc, PhD Candidate1,2, Shahram Lak; PhD2, Mani Mojaddam; PhD2*,Ahmad Naderi; PhD2,3 & Mohammad Abdollahian-Noghabi; PhD2,4
Sina Fallah-Tafty; MSc, PhD Candidate1,2, Shahram Lak; PhD2, Mani Mojaddam; PhD2*,Ahmad Naderi; PhD2,3 & Mohammad Abdollahian-Noghabi; PhD2,4
1 Department of Agronomy, Khuzestan Science and Research Branch, Islamic Azad University, Ahvaz, Iran.
2 Department of Agronomy, Ahvaz Branch, Islamic Azad University, Ahvaz, Iran.
3 Seed and Plant Improvement Institute, Agricultural Research, Education and Extension Organization, Ahvaz, Iran.
4 Agricultural Research, Education and Extension Organization, Tehran, Iran.
| ARTICLE INFO | ABSTRACT | |
| ORIGINAL ARTICLE |
|
|
| Article history: Received: 2 Dec 2019 Revised: 22 Jun 2020 Accepted: 29 Mar 2020 |
||
| *Corresponding author: manimojaddam@yahoo.com Department of Agronomy, Ahvaz Branch, Islamic Azad University, Ahvaz, Iran. Postal code:3143844495 Tel: +98-9166124437 |
Introduction
Soybean is the main legume and oil seed crop across the world that contains the main sources of animal protein food and is important for food chain production (Argaw, 2012). During food production, weeds and pest interference will reduce yield. Among numerous ways to control weeds, using chemicals are the easiest ones, but they are highly dangerous for the environment and human beings. Recently, researchers used many ways to reduce herbicide wastes and residues in the environment, but tank-mixed and reduced herbicide rates were the most efficient ones (Barroso et al., 2010). Long-term application of herbicides will induce herbicide resistant in weeds, which constraints food production and leads to poor quality and quantity productions (Délye et al., 2013, Heap, 2014, Yuan et al., 2007).Tank-mixed herbicides increase poison mode of action and enhance weeds control by changing herbicides’ active ingredient (Hatzio and Penner, 1985, Zhang et al., 1995). Furthermore, tank-mixed herbicide particles interact with each other in many synergic, additive, and antagonistic ways. When synergic effect is detected, reduction of the herbicide rate is recommendable. This advantage provides the farmers with the opportunity to reduce herbicide rates due to economic, technical, and environmental reasons. As a result, the poison drift and residue are reduced in the final productions, which guarantee sustainable food chain (Blackshaw et al., 2006), Pannacci and Covarelli, 2009). Imazethapyr prohibits grass and broad leaf weeds’ growth by inhibiting amino acid synthesis and accumulating in the meristemic plant body parts, which finally inhibit the plant growth. Imazethapyr recommended rate for soybean field is 100 g active ingredisents/hectare (g ai/ha) (Krausz et al., 2001). Bentazon is a post emergence herbicide that exposes the plants to bentazon damaged by reactive oxygen species (ROS) and affects protein and cell membrane (Ahrens, 1994, Armel et al., 2007, Hugie et al., 2008, Powles and Yu, 2010). Bentazon activity is in chloroplast of broad-leafed weeds and its recommended rate for soybean field is 960 g ai/ha. (Han and Wang, 2002, Williams and Nelson, 2014, Zhu et al., 2009). Sethoxydim, a post emergence systemic herbicide, impacts grass weeds’ infestation within lipid synthesis inhibition mode of action and Sethoxydim recommended rate for soybean field is 375 g ai/ha. Tank-mixed herbicide improves weed control by manipulating the site of action and preventing the weed tolerance occurrence. Mixed herbicide modifies the enzymes’ activity considering various sites of action. However, single herbicides trigger different enzyme activity and have different effects on soybean. They also detoxify the effects of herbicides in many ways such as closing stomatal as well as using hormones and antioxidant enzymes (Alexieva et al., 2001, Caverzan et al., 2016, Czarnocka and Karpiński, 2018, Mittler, 2002). Moreover, closed stomatal is in the front line of herbicide uptake prevention which induces accumulation of ROS produced by unused energy (Boulahia et al., 2016, Jiang and Yang., 2009, Pan et al., 2017, Zhang et al., 2014). By closing stomatal, Carbon dioxide cannot take apart in photosynthesis since sunlight is absorbed and electron chain continues working dramatically towards ROS production. Finally, these reactions can damage the cell wall, DNA, and proteins (Bailly, 2004, Bailly et al., 1996, Foyer and Noctor, 2005, Mittler, 2017, Mühling and Läuchli, 2003, Tan et al., 2006, Xu et al., 2010, Yordanova et al., 2004). Superoxide (O2) is the most dangerous ROS that harms cells in plants due to high oxidative ability (Jung, 2004, Triantaphylidès and Michel, 2009). Superoxide dismutase (SOD) catalyzes superoxide anion to hydrogen peroxide (H2O2) (Babior et al., 1975, Galeshi et al., 2009, Gill and Tuteja, 2010, Li et al., 2014), where Ascorbate peroxidase (APX) (Kafi et al., 2009, Wang et al., 2004), Glutathione reductase (GR) (Ahmed et al., 2002), Catalase (Airam et al., 2009, Dubey, 2010), and Dehydro ascorbate reductase (DHAR) (Anjum et al., 2014, Gupta et al., 2001) transform H2O2 to H2O and O2. Activity of these 5 enzymes revealed soybean ability against side effects of herbicides and proposed its detoxification ability in consuming the energy of final yield that minimized the herbicides’ residue in yield and reduced the environment harms. So, these antioxidant enzymes play the main role in producing safe foods during soybean production.
The aim of this study was to find the best herbicide component to promote minimum enzyme activity affecting food quantity and quality directly. Moreover, we aimed to characterize the best reduced herbicide dose against weeds’ infestation.
Materials and Methods
Treatments characteristics: This study was carried out by DPX cultivar of soybean planted in ploughed field at different plots during 2014 in growing seasons. The experiments were carried out in two locations of Alborz province including a research farms in Islamic Azad University of Karaj and Sugar Beet Research Farm Institute at Kamalshahr. Each experimental unit had an area of about 18 m2, which included 6 soybean rows with 50 cm lateral distance. Weed flora was different in each area showing a wide aspect of herbicide usage that affects yield. The soil texture in the first and second locations were loamy sandy and sandy loamy, respectively. This study composed of two main treatments, which include herbicides in 7 levels of single usage of imazethapye, bentazon and sethoxydim, double solution of imazethapyr + bentazon, imazethapyr + sethoxydim and bentazon + sethoxydim, and finally triple solution of imazethapyr + bentazon + sethoxydim as one solution. The second treatment included different herbicide rates containing the full recommended dosage of 100%, reduced to 60% and 30% of the label recommended dose. Exact dose calculation of each herbicide is proposed in Table1. These treatments were applied at leaf expanding growth stage of soybean by a backpack sprayer using flat nuzzle.
Sampling assay: Soybean yield was harvested 95 days after planting and the collected samples were weighted. To survey the enzymes’ activity, leaf samples were collected 3 days after herbicide application. Immediately, they were frozen by liquid nitrogen. Later, the samples were extracted by a pestle from the ice cooled crasher using 4 ml of 0.05 M Na2Hpo4/NaH2Po4 (pH 7.0) buffer containing 0.2 µM Ethylene Diamine Tetracetic Acid (EDTA) and 1% Poly Vinil-Pyrolidone (PVP). The homogenates were centrifuged at 4oC for 20 min at 15000 rpm (Zhang et al., 2005). Supernatants were collected and used for enzymes’ activity assay.
Ascorbate peroxidase (APX) assay: Ascorbate peroxidase activity was measured according to Nakano and Asada procedure, which depended on decreasing absorbance at 290 nm, while the ascorbate was oxidized. Reaction mixture composed of 50 µM/l Na-phosphate buffer (pH 7.0), 50 µM/l ascorbate, 0.1 µM/l EDTA, 1.2 µM/l H2O2, and 0.1 ml of enzyme extract in a final assay volume of 1 ml. Concentration of oxidized ascorbate was calculated by coefficient of 2.8 µM/l/cm. Each unit of APX included reduction of 1 µM/ml/min ascorbate oxidized (Nakano and Kozi, 1981).
Catalase (CAT) assay: CAT extract (20 ml), was added to reaction component, which included 750 ml hydrogen peroxidase (H2O2) and 750 ml of 100 µM phosphate buffer (pH 7.0). Later, this solution was adjusted to 3 ml with sterile distilled water. Finally, the absorbance was read at 240 nm.
Glutathione reductase (GR) assay: GR activity was measured as Foyer and Halliwell assay. Container consisted of 25 µM Na-phosphate buffer (pH 7.8), 0.5 µM GSSG, 0.12 µM NADPH, and 0.1 ml enzyme extract in a final assay volume of 1 ml. NADPH oxidation intercepted at 340 nm. Activity was calculated with extinction coefficient of NADPH (6.2 µM/cm). Each unit of GR included reduction of 1 µM/ml/min glutathione (Foyer and Halliwell, 1976).
Dehydroascorbate Reductase (DHAR) assay: DHAR was measured by reducing 0.7 ml phosphate buffer (pH 7.0), 20 µM/l of reduced glutathione (GSH) in phosphate buffer (pH 7.0), 2 µM/l DHA, and 0.1 ml crude enzyme. Freshly prepared DHAR, kept on ice, was added to the reaction mixture in covette. Reduction of DHAR to ASA was monitored by increase of absorbance at 290 nm, taking 2.8 µM/l/cm as the absorbance coefficient (Krivosheeva et al., 1996).
Superoxide dismutase (SOD) assay: Activity of SOD was calculated by the photoreduction of Nitortetrazolium Blue Chloride (NBT). Reaction solution contained 100 µM phosphate buffer
(pH 7.0), 0.1 µM EDTA, 13 µM methionine,
75 µM Nitrotetrazolium Blue Choloride, 2 µM riboflavin, and adequate supernatant. Riboflavin was added to solution as lazy component and then the reaction started by a 15 watts’ florescent lamp. After the reaction ended, the reaction product was measured at 560 nm. Isoenzymes of SOD were separated on 10% none-denaturing PAGE at 4°C. Finally, the same volume of each sample was loaded to this solution. These extracts were electrophoresed and SOD activity was calculated by monitoring according to Demirevska-Kepova procedure (Demirevska-Kepova et al., 2004).
Data analysis: Data obtained by different methods were analyzed by SAS software to determine the treatment effect on enzymes’ activity. Completely randomized factorial design was used with three replications to analyze data variance and to determine its significant treatment effect on yield and enzymes. To investigate the effect of both treatments (herbicide and doses) on enzymes, the mean comparisons’ method was used based on DUNCAN procedure.
Results
Analysis of variance revealed that both main treatments (herbicide rates and herbicide) had significant effect on soybean yield. In other words, antioxidant activity varied by changing herbicides at different rates and affected the final yield (Table 2). The maximum soybean yield was registered at tank-mixed Imazethapyr + Bentazon + Sethoxydim treatment within all rates. The findings showed that the maximum rate was 3.7 ton/hectare (t/ha) at full recommended rate of 100% (including 33 g Imazethapyr + 320 g Bentazon + 125 g Sethoxydim active ingredients as a one solution) and 3.1 t/ha when herbicide rate was reduced to 30% of the label recommended rate (which included 10 g Imazethapyr + 96 g Bentazon + 33 g Sethoxydim active ingredients as one solution) (Table 3). Minimum yield registered during single herbicide treatment was 2.2 t/ha for imazethapyr, 2 t/ha for bentazon, and 1.2 t/ha for Sethoxydim (Table 4). The maximum activity of enzymes registered at all herbicide treatments induced a full rate of 100%, where the minimum enzymes’ activity was demonstrated at minimum herbicide rate of 30% (Table 5) during tank mixed treatments (Table 4). All enzymes’ activity units were international unit in one gram of the sample international unit/gram (iu/g) (which was explained at material and methods’ section) and soybean yield was t/ha.
The aim of this study was to find the best herbicide component to promote minimum enzyme activity affecting food quantity and quality directly. Moreover, we aimed to characterize the best reduced herbicide dose against weeds’ infestation.
Materials and Methods
Treatments characteristics: This study was carried out by DPX cultivar of soybean planted in ploughed field at different plots during 2014 in growing seasons. The experiments were carried out in two locations of Alborz province including a research farms in Islamic Azad University of Karaj and Sugar Beet Research Farm Institute at Kamalshahr. Each experimental unit had an area of about 18 m2, which included 6 soybean rows with 50 cm lateral distance. Weed flora was different in each area showing a wide aspect of herbicide usage that affects yield. The soil texture in the first and second locations were loamy sandy and sandy loamy, respectively. This study composed of two main treatments, which include herbicides in 7 levels of single usage of imazethapye, bentazon and sethoxydim, double solution of imazethapyr + bentazon, imazethapyr + sethoxydim and bentazon + sethoxydim, and finally triple solution of imazethapyr + bentazon + sethoxydim as one solution. The second treatment included different herbicide rates containing the full recommended dosage of 100%, reduced to 60% and 30% of the label recommended dose. Exact dose calculation of each herbicide is proposed in Table1. These treatments were applied at leaf expanding growth stage of soybean by a backpack sprayer using flat nuzzle.
Sampling assay: Soybean yield was harvested 95 days after planting and the collected samples were weighted. To survey the enzymes’ activity, leaf samples were collected 3 days after herbicide application. Immediately, they were frozen by liquid nitrogen. Later, the samples were extracted by a pestle from the ice cooled crasher using 4 ml of 0.05 M Na2Hpo4/NaH2Po4 (pH 7.0) buffer containing 0.2 µM Ethylene Diamine Tetracetic Acid (EDTA) and 1% Poly Vinil-Pyrolidone (PVP). The homogenates were centrifuged at 4oC for 20 min at 15000 rpm (Zhang et al., 2005). Supernatants were collected and used for enzymes’ activity assay.
Ascorbate peroxidase (APX) assay: Ascorbate peroxidase activity was measured according to Nakano and Asada procedure, which depended on decreasing absorbance at 290 nm, while the ascorbate was oxidized. Reaction mixture composed of 50 µM/l Na-phosphate buffer (pH 7.0), 50 µM/l ascorbate, 0.1 µM/l EDTA, 1.2 µM/l H2O2, and 0.1 ml of enzyme extract in a final assay volume of 1 ml. Concentration of oxidized ascorbate was calculated by coefficient of 2.8 µM/l/cm. Each unit of APX included reduction of 1 µM/ml/min ascorbate oxidized (Nakano and Kozi, 1981).
Catalase (CAT) assay: CAT extract (20 ml), was added to reaction component, which included 750 ml hydrogen peroxidase (H2O2) and 750 ml of 100 µM phosphate buffer (pH 7.0). Later, this solution was adjusted to 3 ml with sterile distilled water. Finally, the absorbance was read at 240 nm.
Glutathione reductase (GR) assay: GR activity was measured as Foyer and Halliwell assay. Container consisted of 25 µM Na-phosphate buffer (pH 7.8), 0.5 µM GSSG, 0.12 µM NADPH, and 0.1 ml enzyme extract in a final assay volume of 1 ml. NADPH oxidation intercepted at 340 nm. Activity was calculated with extinction coefficient of NADPH (6.2 µM/cm). Each unit of GR included reduction of 1 µM/ml/min glutathione (Foyer and Halliwell, 1976).
Dehydroascorbate Reductase (DHAR) assay: DHAR was measured by reducing 0.7 ml phosphate buffer (pH 7.0), 20 µM/l of reduced glutathione (GSH) in phosphate buffer (pH 7.0), 2 µM/l DHA, and 0.1 ml crude enzyme. Freshly prepared DHAR, kept on ice, was added to the reaction mixture in covette. Reduction of DHAR to ASA was monitored by increase of absorbance at 290 nm, taking 2.8 µM/l/cm as the absorbance coefficient (Krivosheeva et al., 1996).
Superoxide dismutase (SOD) assay: Activity of SOD was calculated by the photoreduction of Nitortetrazolium Blue Chloride (NBT). Reaction solution contained 100 µM phosphate buffer
(pH 7.0), 0.1 µM EDTA, 13 µM methionine,
75 µM Nitrotetrazolium Blue Choloride, 2 µM riboflavin, and adequate supernatant. Riboflavin was added to solution as lazy component and then the reaction started by a 15 watts’ florescent lamp. After the reaction ended, the reaction product was measured at 560 nm. Isoenzymes of SOD were separated on 10% none-denaturing PAGE at 4°C. Finally, the same volume of each sample was loaded to this solution. These extracts were electrophoresed and SOD activity was calculated by monitoring according to Demirevska-Kepova procedure (Demirevska-Kepova et al., 2004).
Data analysis: Data obtained by different methods were analyzed by SAS software to determine the treatment effect on enzymes’ activity. Completely randomized factorial design was used with three replications to analyze data variance and to determine its significant treatment effect on yield and enzymes. To investigate the effect of both treatments (herbicide and doses) on enzymes, the mean comparisons’ method was used based on DUNCAN procedure.
Results
Analysis of variance revealed that both main treatments (herbicide rates and herbicide) had significant effect on soybean yield. In other words, antioxidant activity varied by changing herbicides at different rates and affected the final yield (Table 2). The maximum soybean yield was registered at tank-mixed Imazethapyr + Bentazon + Sethoxydim treatment within all rates. The findings showed that the maximum rate was 3.7 ton/hectare (t/ha) at full recommended rate of 100% (including 33 g Imazethapyr + 320 g Bentazon + 125 g Sethoxydim active ingredients as a one solution) and 3.1 t/ha when herbicide rate was reduced to 30% of the label recommended rate (which included 10 g Imazethapyr + 96 g Bentazon + 33 g Sethoxydim active ingredients as one solution) (Table 3). Minimum yield registered during single herbicide treatment was 2.2 t/ha for imazethapyr, 2 t/ha for bentazon, and 1.2 t/ha for Sethoxydim (Table 4). The maximum activity of enzymes registered at all herbicide treatments induced a full rate of 100%, where the minimum enzymes’ activity was demonstrated at minimum herbicide rate of 30% (Table 5) during tank mixed treatments (Table 4). All enzymes’ activity units were international unit in one gram of the sample international unit/gram (iu/g) (which was explained at material and methods’ section) and soybean yield was t/ha.
| Table 1. Single and tank-mixed herbicide treatment rates calculations | ||||||||||||
| Active ingredients rate |
Treatments ( Herbicide & Herbicide Rates) | |||||||||||
| IBS100 | IBS60 | IBS30 | IB100 | IB60 | IB30 | IS100 | IS60 | IS30 | BS100 | |||
| Imazethapyr (g) | 33 | 20 | 10 | 50 | 30 | 15 | 50 | 30 | 15 | 480 | ||
| Betazon (g) | 320 | 192 | 96 | 480 | 288 | 144 | 187 | 112 | 56 | 187 | ||
| Sethoxydim (g) | 125 | 75 | 37 | - | - | - | - | - | - | - | ||
| Active ingredients rate | BS60 | BS30 | I100 | I60 | I30 | B100 | B60 | B30 | S100 | S60 | S30 | |
| Imazethapyr (g) | 288 | 144 | 100 | 60 | 30 | 960 | 576 | 288 | 375 | 225 | 112 | |
| Betazon (g) | 112 | 56 | - | - | - | - | - | - | - | - | - | |
| Sethoxydim (g) | - | - | - | - | - | - | - | - | - | - | - | |
| IB = Imazethapyr + Bentazon, IS = Imazethapyt + Sethoxydim, BS = Bentazon + Sethoxydim, IBS = Imazethapyr + Bentazon + Sethoxydim, 100 = Full recommended rate of herbicide, 60= Reduced to 60 percent of the label recommended rate, 30 = Reduced to 30 percent of label recommended rate. | ||||||||||||
| Table 2. Analysis of variances of soybean yield and enzymes activity (SOD, APX, CAT, GR and DHAR) | |||||||
| Source of variance | DF | Yield | SOD | APX | CAT | GR | DHAR |
| Location | 1 | 0.24a | 0.31a | 0.30 a | 0.36 a | 0.30 a | 0.11a |
| Block×Location | 4 | 0.04a | 0.44a | 0.40a | 0.62a | 0.19a | 0.03a |
| Herbicide | 6 | 10.9a | 33.6a | 14.5a | 18.0 a | 5.4a | 0.8a |
| Herbicide dose | 2 | 5.86a | 105.0a | 17.4a | 8.0a | 2.1a | 0.3a |
| Herbicide×Herbicide dose | 12 | 0.15a | 1.1a | 0.7a | 0.07a | 0.04a | 0.001a |
| Location×Herbicide | 6 | 0.01b | 0 b | 0.001b | 0b | 0b | 0b |
| Location×Dose | 2 | 0.01b | 0b | 0b | 0b | 0b | 0b |
| Location×Herbicide×Dose | 12 | 0.008b | 0b | 0.001b | 0b | 0b | 0b |
| Error | 80 | 0.009 | 0.001 | 0.001 | 0.001 | 0.009 | 0.0001 |
| CV | 3.6 | 0.8 | 0.92 | 0.99 | 3 | 2.3 | |
| a: significant at 5% and 1% probability level, b: Non-significant, SOD: Superoxide dismutase, APX: Ascorbate peroxidase, CAT: Catalase, GR: Glutathione reductase , DHAR:Dehydroascorbate Reductase | |||||||
| Table 3. Mean of both location by induction treatment on soybean yield and enzym activity (SOD, APX, CAT, GR and DHAR) | |||||||||||
| Variables | Treatments ( Herbicide & Herbicide Rates) | ||||||||||
| Enzymes activity (iu) | IBS100 | IBS60 | IBS30 | IB100 | IB60 | IB30 | IS100 | IS60 | IS30 | BS100 | BS60 |
| Superoxide dismutase | 3.9 | 3.5 | 1.9 | 6.1 | 5.1 | 3.1 | 5.2 | 4.5 | 2.7 | 4.7 | 4.1 |
| Ascorbate peroxidase | 3.9 | 3.7 | 3.5 | 4..1 | 3.8 | 3.6 | 4.7 | 4.1 | 3.7 | 5.1 | 4.5 |
| Catalase | 2.4 | 2 | 1.7 | 2.7 | 2.2 | 1.9 | 3.1 | 2.6 | 2.4 | 3.5 | 3 |
| Glutathione reductase | 0.4 | 0.3 | 0.2 | 0.6 | 0.5 | 0.3 | 0.8 | 0.6 | 0.4 | 1.1 | 0.9 |
| Dehydroascorbate Reductase | 0.2 | 0.1 | 0.07 | 0.3 | 0.2 | 0.1 | 0.4 | 0.3 | 0.2 | 0.5 | 0.4 |
| Soybean yield (t/ha) | 3.7 | 3.5 | 3.1 | 3.6 | 3.3 | 2.8 | 3.2 | 2.7 | 2.4 | 3.0 | 2.9 |
| Anzymes activity (iu) | BS30 | I100 | I60 | I30 | B100 | B60 | B30 | S100 | S60 | S30 | Control |
| Superoxide dismutase | 2.2 | 8.8 | 6.9 | 4.7 | 7.5 | 5.7 | 3.7 | 8.1 | 6.4 | 4.1 | 0.50 |
| Ascorbate peroxidase | 3.9 | 5.9 | 5.1 | 4.2 | 7.2 | 6 | 4.9 | 6.5 | 5.7 | 4.6 | 1.20 |
| Catalase | 2.7 | 4.2 | 3.9 | 3.2 | 4.7 | 4.3 | 3.7 | 5.2 | 4.8 | 4.1 | 0.50 |
| Glutathione reductase | 0.6 | 1.4 | 1.1 | 0.8 | 2.1 | 1.8 | 1.5 | 1.8 | 1.5 | 1.2 | 0.15 |
| Dehydroascorbate Reductase | 0.3 | 0.6 | 0.5 | 0.4 | 0.8 | 0.7 | 0.6 | 0.7 | 0.6 | 0.5 | 0.02 |
| Soybean yield (t/ha) | 2.7 | 2.7 | 2.3 | 1.7 | 2.6 | 2.1 | 1.4 | 1.7 | 1.1 | 0.7 | 3.8 |
| IB = Imazethapyr + Bentazon, IS = Imazethapyt + Sethoxydim, BS = Bentazon + Sethoxydim, IBS = Imazethapyr + Bentazon + Sethoxydim, 100 = Full recommended rate of herbicide, 60= Reduced to 60 percent of the label recommended rate, 30 = Reduced to 30 percent of label recommended rate. | |||||||||||
| Table 4. Mean comparison of the main effect of herbicide treatment on soybean yield and enzymes activity (SOD, APX, CAT, GR, and DHAR) | ||||||
| Herbicide Treatment | Soybean yield (t/ha) and antioxidant enzymes activity (i.u)a | |||||
| Yield | SOD | APX | CAT | GR | DHAR | |
| Imazethapyr + Bentazon + Sethoxydim | 3.5 A | 3.1 G | 3.7 G | 2.1 G | 0.35 G | 0.15 G |
| Imazethapyr + Bentazon | 3.2 B | 4.8 D | 3.8 F | 2.3 F | 0.51 F | 0.22 F |
| Imazethapyr + Sethoxydim | 2.8 C | 4.1 E | 4.2 E | 2.7 E | 0.65 E | 0.32 E |
| Bentazon + Sethoxydim | 2.8 C | 3.7 F | 4.5 D | 3.1 D | 0.91 D | 0.43 D |
| Imazethapyr | 2.2 D | 6.8 A | 5.1 C | 3.8 C | 1.1 C | 0.53 C |
| Bentazon | 2 E | 5.6 C | 6.1 A | 4.2 B | 1.8 A | 0.72 A |
| Sethoxydim | 1.2 F | 6.2 B | 5.6 B | 4.7 A | 1.5 B | 0.63 B |
| a: Within each column, means with the same letter do not have difference and are grouped as one bunch according to the Duncan test (P≤ 0.05), SOD: Superoxide dismutase , APX: Ascorbate peroxidase , CAT: Catalase , GR: Glutathione reductase , DHAR: Dehydroascorbate Reductase. | ||||||
| Table 5. Mean comparison of the main effect of herbicide doses on soybean yield, enzymes activity (SOD, APX, CAT, GR, and DHAR) . |
||||||
| Herbicide dose | Soybean yield (t/ha) and antioxidant enzymes activity (iu)a | |||||
| Yield | SOD | APX | CAT | GR | DHAR | |
| 100% | 2.9 A | 6.3 A | 5.3 A | 3.7 A | 1.2 A | 0.52 A |
| 60% | 2.6 B | 5.2 B | 4.7 B | 3.3 B | 1.0 B | 0.43 B |
| 30% | 2.2 C | 3.2 C | 4.1 C | 2.8 C | 0.76 C | 0.33 C |
| a: Within each column, means with the same letter do not have difference and are grouped as one bunch according to the Duncan test (P≤ 0.05), SOD: Superoxide dismutase , APX: Ascorbate peroxidase , CAT: Catalase , GR: Glutathione reductase , DHAR: Dehydroascorbate Reductase. | ||||||
| Figure 1. Interaction of herbicides and doses on soybean yield |
| Figure 2. Effect of different herbicides on SOD, APX activity (A) and DHAR, CAT activity (B) |
Figure 3. Effect of different herbicide doses on CAT and DHAR activity
Discussion
Many researchers confirmed constant soybean yield while tank-mixed herbicide was used with lower doses (Auskalnis and Kadzys, 2006, Barros et al., 2005, Boström and Håkan, 2002, Walker et al., 2002, Zhang et al., 2000). However, we are faced with scarcity of information regarding soybean production using herbicide tank-mixing (Imazethapyr, Bentazon and Sethoxidym). This demonstrates reduction of imazethapyr rates by 23 g ai/ha (from 33 g ai/ha in full rate to 10 g ai/ha in the reduced rate), bentazon by 224 g ai/ha (from 320 g ai/ha in full rate to 96 g ai/ha in reduced rate), and sethoxydim by 88 g ai/ha (from 125 g ai/ha in full rate to 37 g ai/ha in reduced rate). When they are used as one tank-mixed solution, herbicide synergic effect will occur and weed control spectrum will increase (due to the manipulating herbicide sites of action that increase soybean yield besides less herbicide utilization) (Table 3). Simultaneous increase of herbicide rates raised enzymes’ activity in all 5 antioxidants (Table 5), which was also confirmed by another researcher (Merve and Burcu, 2012). Long-term application of full rates of single herbicides will consume soybean energy and affect the yield negatively. Many researchers confirm this hypothesis (Boulahia et al., 2016, Jiang and Yang., 2009, Pan et al., 2017, Zhang et al., 2014). For instance, tank-mixed herbicide superoxide dismutase activity reduced 2 iu (from 3.9 iu to 1.9 iu) when imazethapy + bentazon + sethoxydim rate diminished from 100% to 30%. This findings was also confirmed by some researchers saying that reducing herbicide rate will minimized the SOD activity (Alexieva et al., 2001, Caverzan et al., 2016, Czarnocka and Karpiński, 2018, Mittler, 2002). This result confirms our former hypothesis proposing that the lower enzymes’ activity leads to higher soybean yield. In contrast, when single herbicides were used, the minimum soybean yield was registered (when sethoxidym doses was 375 g ai/ha soybean yield reduced to 1.7 t/ha). During this single component treatment, maximum fluctuation was observed in soybean yield; it reduced from 1.7 t/ha at 375 g ai/ha sethoxydim rate to 0.7 t/ha at 112 g ai/ha at reduced rate. This result reported by Rosales revealed that single component herbicide induced the maximum yield fluctuation(Rosales-Robles et al., 2005). Besides, the highest antioxidant enzymes activity was registered at single component herbicide usage (including imazethapyr, bentazon, and sethoxydim). This incident is due to the effect of high active ingredients of each herbicide, when used as single components (Table 1), on soybean. It finally promotes soybean antioxidant enzyme system to eliminate herbicide side effects. For instance, SOD activity rose from 3.1 iu at triple tank-mixed herbicide to 6.2 iu, when sethoxydim was applied as a single herbicide (Table 4). Each enzyme participates in specific herbicide usage; for instance, APX maximum activity was at bentazon (Figure 2A) usage, which was due to the same work place of APX and bentazon (chloroplast and mitochondria). Simultaneously, the highest CAT activity (4.7 iu) and soybean yield reduction (1.2 t/ha in comparison to weed free treatment) (Table 4) were observed at single bentazon usage (269 g ai/ha) (Figure 2B). Moreover, the maximum GR activity (2.1 iu) and soybean yield reduction (1.3 t/ha) (Table 3) were seen at single usage of bentazon (260 g ai/ha). These results confirm the hypothesis saying that herbicides’ treatment with one site of action increases antioxidant enzymes’ activity that consumes the soybean energy and reduces its yield. As a result, single mode of action herbicides put a lot of stress on plant, which can have negative effects on yield and food chain security. This results were confirmed by a study(Knežević et al., 2003). During doubled component herbicide usage, the yield raised in comparison to the single mode of action herbicide, but it was lower compared with the triplet mode of action herbicide. For instance, when Imazethapyr + Bentazon, imazethapyr + Sethoxydim, and bentazon + Sethoxydim solutions induced soybean, the yield raised to 3.2, 2.8, and 2.8 t/ha, respectively in comparison to single herbicides (Table 4 and Figure 1). During yield survey among double component herbicide solution, the maximum soybean yield was registered at Imazethapyr+Bnetazon (Figure 1), where minimum enzymes’ activity occurred was 6.1 iu for SOD,4.1 iu for APX, 2.7 iu (Figure 2A) for CAT, 0.6 iu for GR, and 0.3 iu for DHAR (Table 4 and Figure 2B). Herbicide reduced rates during doubled component herbicide solution had more positive effects on yield but were not reliable in single component herbicide solutions since yield was unstable due to the higher activity of enzymes (Figure1) (Dogan, 2005, Fanadzo et al., 2010). It was also proposed that herbicide rates were more reliable during tank mixed herbicide induction. In contrast, the maximum yield stability was observed in reduced herbicide rates of Imazethapyr + Bentazon + Sethoxydim treatment. In the following, Imazethapyr + Bentazon (at full recommended rate of 50 gr ai/ha Imazethapyr and 480 g ai/ha Bentazon) was ranked the second with regard to the best yield record of reduced herbicide rates in which the soybean yield was 3.7 t/ha. Results also showed that reducing herbicide rate to 30% of the lable recommended soybean yield remained as 3 t/ha (Walker et al., 2002) (Table 3 and Figure 1). The imazethapy + bentazon + sethoxydim and Imazethapyr + Bentazon can be recommended to reduce the usage rate with secure yield besides the minimum herbicide residue in yield which guarantee the food chain health.
Conclusion
According to the findings, tank-mixed herbicides trigger less ROS that lowers priority of soybean antioxidant enzyme activity and increases the yield. It also reduces herbicide residue, which guarantees secure food production. Moreover, herbicides rates can be reduced and fixed yield quantity and quality can be guaranteed by manipulating modes of actions proposed by tank-mixed herbicides. As a result, sustainable secure food production programs can be designed and poison erosion can be prevented.
Author contributions
Fallah-Tafty S and Lak S designed the study; Fallah-Tafty S and Mojaddam M conducted the experiments. Fallah-Tafty S, Abdollahian-Noghabi M, and Naderi A analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Conflicts of interest
All authors declared no conflict of interests.
Acknowledgements
The authors thank all the individuals who were directly or indirectly associated with this experiment. This research was based on a PhD thesis with 23250208972001 ID number conducted in Department of Agronomy, Islamic Azad University of Khouzestan, Science and Research Branch, Ahvaz, Iran.
References
Ahmed S, Nawata E, Hosokawa M, Domae Y & Sakuratani T 2002. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Science 163(1): 117-123.
Ahrens WH 1994. Herbicide hand book.
Airam R, Dharmar K, Chinnusamy V & Meena R 2009. Waterlogging-induced increase in sugar mobilization, fermentation, and related gene expression in the roots of mung bean (Vigna radiata). Journal of Plant Physiology 166(6): 602-616.
Alexieva V, Sergiev I, Mapelli S & E K 2001. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant, Cell & Environment 24(12): 1337-1344.
Anjum N, et al. 2014. Hasanuzzaman, Armando C. Duarte, Eduarda Pereira, Iqbal Ahmad, Renu Tuteja, and Narendra Tuteja. "Metal/metalloid stress tolerance in plants: role of ascorbate, its redox couple, and associated enzymes. Protoplasma.251(6): 1265-1283.
Argaw A 2012. Evaluation of co-inoculation of Bradyrhizobium japonicum and phosphate solubilizing Pseudomonas spp. effect on soybean (Glycine max L.(Merr.)) in Assossa area. Journal of Agricultural Science and Technology 14(1): 213-224.
Armel G, Rardon P, McComrick M & Ferry N 2007. Differential response of several carotenoid biosynthesis inhibitors in mixtures with atrazine. Weed Technology 21(4): 947-953.
Auskalnis A & Kadzys A 2006. Effect of timing and dosage in herbicide application on weed biomass in spring wheat. Agronomy Research 4:133-136.
Babior B, Curnutte J & Kipnes R 1975. Biological defense mechanisms. Evidence for the participation of superoxide in bacterial killing by xanthine oxidase. Journal of Laboratory and Clinical Medicine.85(2): 235-244.
Bailly C 2004. Active oxygen species and antioxidants in seed biology. Seed Science Research.14(2): 93-107.
Bailly C, Benamar A, Corbineau F & Côme D 1996. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiologia Plantarum.97(1): 104-110.
Barros J, Basch G & de Carvalho M 2005. Effect of reduced doses of a post-emergence graminicide mixture to control Lolium rigidum G. in winter wheat under direct drilling in Mediterranean environment. Crop Protection.24(10): 880-887.
Barroso A, Yamauti M & Alves P 2010. Interference between weed species and two bean cultivars in two times of sowing. Bragantia.69(3): 609-616.
Blackshaw R, Odonovan J, Harker K, Clayton G & Stougaard R 2006. Reduced herbicide doses in field crops: a review. Weed Biology and Management.6(1): 10-17.
Boström U & Håkan F 2002. Response of weeds and crop yield to herbicide dose decision-support guidelines. Weed Science 50, no. 2 50(2): 186-195.
Boulahia K, Carol P, Planchais S & Abrous-Belbachir O 2016. Phaseolus vulgaris L. seedlings exposed to prometryn herbicide contaminated soil trigger an oxidative stress response. Journal of Agricultural and Food Chemistry 64(16): 3150-3160.
Caverzan A, Casassola A & Brammer S 2016. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. SHANKER AK & SHANKER C. Abiotic and biotic stress in plants-Recent advances and future perspectives. In-Tech.
Czarnocka W & Karpiński S 2018. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radical Biology and Medicine 122: 4-20.
Délye C, Jasieniuk M & Le Corre V 2013. Deciphering the evolution of herbicide resistance in weeds. Trends in Genetics 29(11): 649-658.
Demirevska-Kepova K, Simova-Stoilova L, Stoyanova Z, Hölzer R & Feller U 2004. Biochemical changes in barley plants after excessive supply of copper and manganese. Environmental and Experimental Botany 52, no. 3 52(3): 253-266.
Dogan M 2005. Efficacies of reduced herbicide rates for weed control in maize (Zea mays L.) during critical period. Journal of Agronomy 4(1): 44-48.
Dubey R 2010. Metal toxicity, oxidative stress and antioxidative defense system in plants. Reactive Oxygen Species and Antioxidants in Higher Plants.15: 193-220.
Fanadzo M, Chiduza C & Mnkeni P 2010. Reduced dosages of atrazine and narrow rows can provide adequate weed control in smallholder irrigated maize (Zea mays L.) production in South Africa. African Journal of Biotechnology.9(45): 7660-7666.
Foyer C & Halliwell B 1976. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta.133(1): 21-25.
Foyer C & Noctor G 2005. Redox homeostasis and antioxidant signaling: a metabolic
interface between stress perception and physiological responses. The Plant Cell.17(7): 1866-1875.
Galeshi S, Torabi B, Resam G, RahemiKarizaki A & Barzegar A 2009. Stress management in plants. Publications Gorgan University of Agricultural Sciences and Natural Resources. 24-29.
Gill S & Tuteja N 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry.48(12): 909-930.
Gupta N, Gupta S & Kumar A 2001. Effect of water stress on physiological attributes and their relationship with growth and yield of wheat cultivars at different stages. Journal of Agronomy and Crop Science.186(1): 55-62.
Han Y & Wang C 2002. Physiological basis of bentazon tolerance in rice (Oryza sativa L.) lines. Weed Biology and Management.2(4): 186-193.
Hatzio K & Penner D 1985. Interactions of herbicides with other agrochemicals in higher plants. Reviews of weed science (USA). 5(2): 173-182.
Heap I 2014. Global perspective of herbicide‐resistant weeds. Pest Management Science.70(9): 1306-1315.
Hugie J, Bollero G, Tranel P & Riechers D 2008. Defining the rate requirements for synergism between mesotrione and atrazine in redroot pigweed (Amaranthus retroflexus). Weed Science.56(2): 265-270.
Jiang L & Yang. H 2009. Prometryne-induced oxidative stress and impact on antioxidant enzymes in wheat. Ecotoxicology and Environmental Safety.72(6): 1687-1693.
Jung S 2004. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Science.166(2): 459-466.
Kafi M, et al. 2009. Physiology of environmental stresses in plants, Mashhad: Academic Center for Education, Culture and Research. 87-96.
Knežević M, Đurkić M, Knežević I, Antonić O & Jelaska S 2003. Effects of tillage and reduced herbicide doses on weed biomass production in winter and spring cereals. Plant, Soil and Environment.49(9): 414-421.
Krausz R, Young B, Kapusta G & Matthews J 2001. Influence of weed competition and herbicides on glyphosate-resistant soybean (Glycine max). Weed Technology.15(3): 530-534.
Krivosheeva A, et al. 1996. Cold acclimation and photoinhibition of photosynthesis in Scots pine. Planta.200(3): 296-305.
Li H, et al. 2014. Overexpression of Pp14-3-3 from Pyrus pyrifolia fruit increases drought and salt tolerance in transgenic tobacco plant. Biologia.69(7): 880-887.
Merve A & Burcu S 2012. Changes in physiological parameters and some antioxidant enzymes activities of soybean (Glycine max L. Merr.) leaves under cadmium and salt stress. Journal of Stress Physiology & Biochemistry 8(4).
Mittler R 2002. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science.7(9): 405-410.
Mittler R 2017. Ros are good. Trends in Plant Science.22(1): 11-19.
Mühling K & Läuchli A 2003. Interaction of NaCl and Cd stress on compartmentation pattern of cations, antioxidant enzymes and proteins in leaves of two wheat genotypes differing in salt tolerance. Plant and Soil.253(1): 219-231.
Nakano Y & Kozi A 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant and Cell Physiology 22(5): 867-880.
Pan D, et al. 2017. Interactions between salicylic acid and antioxidant enzymes tilting the balance of H2O2 from photorespiration in non-target crops under halosulfuron-methyl stress. Pesticide Biochemistry and Physiology.143: 214-223.
Powles S & Yu Q 2010. Evolution in action: plants resistant to herbicides. Annual Review of Plant Biology.61: 317-347.
Rosales-Robles E, Sanchez-de-la-Cruz R, Salinas-Garcia J & Pecina-Quintero V 2005. Broadleaf weed management in grain sorghum with reduced rates of postemergence herbicides. Weed Technology 19(2): 385-390.
Tan Y, Zongsuo L, Hongbo S & Feng D 2006. Effect of water deficits on the activity of anti-oxidative enzymes and osmoregulation among three different genotypes of Radix Astragali at seeding stage. Colloids and Surfaces B: Biointerfaces.49(1): 60-65.
Triantaphylidès C & Michel H 2009. Singlet oxygen in plants: production, detoxification and signaling. Trends in Plant Science.14(4): 219-228.
Walker S, Medd R, Robinson G & Cullis B 2002. Improved management of Avena ludoviciana and Phalaris paradoxa with more densely sown wheat and less herbicide. Weed Research 42(4): 257-270.
Wang S, et al. 2004. Copper-induced stress and antioxidative responses in roots of Brassica juncea L. Botanical Bulletin of Academia Sinica.45.
Williams M & Nelson R 2014. Vegetable soybean tolerance to bentazon, fomesafen, imazamox, linuron, and sulfentrazone. Weed Technology.28(4): 601-607.
Xu J, Hengxia Y, Xiaojing L & Xia L 2010. Salt affects plant Cd-stress responses by modulating growth and Cd accumulation. Planta.231(2): 449-459.
Yordanova R, Christov K & Popova L 2004. Antioxidative enzymes in barley plants subjected to soil flooding. Environmental and Experimental Botany.51(2): 93-101.
Yuan J, Tranel P & Stewart Jr C 2007. Non-target-site herbicide resistance: a family business. Trends in plant science.12(1): 6-13.
Zhang J, Hamill A & Weaver S 1995. Antagonism and synergism between herbicides: trends from previous studies. Weed Technology. 86-90.
Zhang J, Lu Y, Zhang J, Tan L & Yang H 2014. Accumulation and toxicological response of atrazine in rice crops. Ecotoxicology and Environmental Safety.102: 105-112.
Zhang J, Weaver S & Hamill A 2000. Risks and reliability of using herbicides at below-labeled rates. Weed Technology.14(1): 106-115.
Zhang X, Ervin E, Evanylo G, Sherony C & Peot C 2005. Biosolids impact on tall fescue drought resistance. Journal of Residuals Science & Technology.2: 173-180.
Zhu J, Patzoldt W, Radwan O, Tranel P & Clough S 2009. Effects of photosystem-II-interfering herbicides atrazine and bentazon on the soybean transcriptome. Plant Genome.2(2): 191-205.
Many researchers confirmed constant soybean yield while tank-mixed herbicide was used with lower doses (Auskalnis and Kadzys, 2006, Barros et al., 2005, Boström and Håkan, 2002, Walker et al., 2002, Zhang et al., 2000). However, we are faced with scarcity of information regarding soybean production using herbicide tank-mixing (Imazethapyr, Bentazon and Sethoxidym). This demonstrates reduction of imazethapyr rates by 23 g ai/ha (from 33 g ai/ha in full rate to 10 g ai/ha in the reduced rate), bentazon by 224 g ai/ha (from 320 g ai/ha in full rate to 96 g ai/ha in reduced rate), and sethoxydim by 88 g ai/ha (from 125 g ai/ha in full rate to 37 g ai/ha in reduced rate). When they are used as one tank-mixed solution, herbicide synergic effect will occur and weed control spectrum will increase (due to the manipulating herbicide sites of action that increase soybean yield besides less herbicide utilization) (Table 3). Simultaneous increase of herbicide rates raised enzymes’ activity in all 5 antioxidants (Table 5), which was also confirmed by another researcher (Merve and Burcu, 2012). Long-term application of full rates of single herbicides will consume soybean energy and affect the yield negatively. Many researchers confirm this hypothesis (Boulahia et al., 2016, Jiang and Yang., 2009, Pan et al., 2017, Zhang et al., 2014). For instance, tank-mixed herbicide superoxide dismutase activity reduced 2 iu (from 3.9 iu to 1.9 iu) when imazethapy + bentazon + sethoxydim rate diminished from 100% to 30%. This findings was also confirmed by some researchers saying that reducing herbicide rate will minimized the SOD activity (Alexieva et al., 2001, Caverzan et al., 2016, Czarnocka and Karpiński, 2018, Mittler, 2002). This result confirms our former hypothesis proposing that the lower enzymes’ activity leads to higher soybean yield. In contrast, when single herbicides were used, the minimum soybean yield was registered (when sethoxidym doses was 375 g ai/ha soybean yield reduced to 1.7 t/ha). During this single component treatment, maximum fluctuation was observed in soybean yield; it reduced from 1.7 t/ha at 375 g ai/ha sethoxydim rate to 0.7 t/ha at 112 g ai/ha at reduced rate. This result reported by Rosales revealed that single component herbicide induced the maximum yield fluctuation(Rosales-Robles et al., 2005). Besides, the highest antioxidant enzymes activity was registered at single component herbicide usage (including imazethapyr, bentazon, and sethoxydim). This incident is due to the effect of high active ingredients of each herbicide, when used as single components (Table 1), on soybean. It finally promotes soybean antioxidant enzyme system to eliminate herbicide side effects. For instance, SOD activity rose from 3.1 iu at triple tank-mixed herbicide to 6.2 iu, when sethoxydim was applied as a single herbicide (Table 4). Each enzyme participates in specific herbicide usage; for instance, APX maximum activity was at bentazon (Figure 2A) usage, which was due to the same work place of APX and bentazon (chloroplast and mitochondria). Simultaneously, the highest CAT activity (4.7 iu) and soybean yield reduction (1.2 t/ha in comparison to weed free treatment) (Table 4) were observed at single bentazon usage (269 g ai/ha) (Figure 2B). Moreover, the maximum GR activity (2.1 iu) and soybean yield reduction (1.3 t/ha) (Table 3) were seen at single usage of bentazon (260 g ai/ha). These results confirm the hypothesis saying that herbicides’ treatment with one site of action increases antioxidant enzymes’ activity that consumes the soybean energy and reduces its yield. As a result, single mode of action herbicides put a lot of stress on plant, which can have negative effects on yield and food chain security. This results were confirmed by a study(Knežević et al., 2003). During doubled component herbicide usage, the yield raised in comparison to the single mode of action herbicide, but it was lower compared with the triplet mode of action herbicide. For instance, when Imazethapyr + Bentazon, imazethapyr + Sethoxydim, and bentazon + Sethoxydim solutions induced soybean, the yield raised to 3.2, 2.8, and 2.8 t/ha, respectively in comparison to single herbicides (Table 4 and Figure 1). During yield survey among double component herbicide solution, the maximum soybean yield was registered at Imazethapyr+Bnetazon (Figure 1), where minimum enzymes’ activity occurred was 6.1 iu for SOD,4.1 iu for APX, 2.7 iu (Figure 2A) for CAT, 0.6 iu for GR, and 0.3 iu for DHAR (Table 4 and Figure 2B). Herbicide reduced rates during doubled component herbicide solution had more positive effects on yield but were not reliable in single component herbicide solutions since yield was unstable due to the higher activity of enzymes (Figure1) (Dogan, 2005, Fanadzo et al., 2010). It was also proposed that herbicide rates were more reliable during tank mixed herbicide induction. In contrast, the maximum yield stability was observed in reduced herbicide rates of Imazethapyr + Bentazon + Sethoxydim treatment. In the following, Imazethapyr + Bentazon (at full recommended rate of 50 gr ai/ha Imazethapyr and 480 g ai/ha Bentazon) was ranked the second with regard to the best yield record of reduced herbicide rates in which the soybean yield was 3.7 t/ha. Results also showed that reducing herbicide rate to 30% of the lable recommended soybean yield remained as 3 t/ha (Walker et al., 2002) (Table 3 and Figure 1). The imazethapy + bentazon + sethoxydim and Imazethapyr + Bentazon can be recommended to reduce the usage rate with secure yield besides the minimum herbicide residue in yield which guarantee the food chain health.
Conclusion
According to the findings, tank-mixed herbicides trigger less ROS that lowers priority of soybean antioxidant enzyme activity and increases the yield. It also reduces herbicide residue, which guarantees secure food production. Moreover, herbicides rates can be reduced and fixed yield quantity and quality can be guaranteed by manipulating modes of actions proposed by tank-mixed herbicides. As a result, sustainable secure food production programs can be designed and poison erosion can be prevented.
Author contributions
Fallah-Tafty S and Lak S designed the study; Fallah-Tafty S and Mojaddam M conducted the experiments. Fallah-Tafty S, Abdollahian-Noghabi M, and Naderi A analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Conflicts of interest
All authors declared no conflict of interests.
Acknowledgements
The authors thank all the individuals who were directly or indirectly associated with this experiment. This research was based on a PhD thesis with 23250208972001 ID number conducted in Department of Agronomy, Islamic Azad University of Khouzestan, Science and Research Branch, Ahvaz, Iran.
References
Ahmed S, Nawata E, Hosokawa M, Domae Y & Sakuratani T 2002. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Science 163(1): 117-123.
Ahrens WH 1994. Herbicide hand book.
Airam R, Dharmar K, Chinnusamy V & Meena R 2009. Waterlogging-induced increase in sugar mobilization, fermentation, and related gene expression in the roots of mung bean (Vigna radiata). Journal of Plant Physiology 166(6): 602-616.
Alexieva V, Sergiev I, Mapelli S & E K 2001. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant, Cell & Environment 24(12): 1337-1344.
Anjum N, et al. 2014. Hasanuzzaman, Armando C. Duarte, Eduarda Pereira, Iqbal Ahmad, Renu Tuteja, and Narendra Tuteja. "Metal/metalloid stress tolerance in plants: role of ascorbate, its redox couple, and associated enzymes. Protoplasma.251(6): 1265-1283.
Argaw A 2012. Evaluation of co-inoculation of Bradyrhizobium japonicum and phosphate solubilizing Pseudomonas spp. effect on soybean (Glycine max L.(Merr.)) in Assossa area. Journal of Agricultural Science and Technology 14(1): 213-224.
Armel G, Rardon P, McComrick M & Ferry N 2007. Differential response of several carotenoid biosynthesis inhibitors in mixtures with atrazine. Weed Technology 21(4): 947-953.
Auskalnis A & Kadzys A 2006. Effect of timing and dosage in herbicide application on weed biomass in spring wheat. Agronomy Research 4:133-136.
Babior B, Curnutte J & Kipnes R 1975. Biological defense mechanisms. Evidence for the participation of superoxide in bacterial killing by xanthine oxidase. Journal of Laboratory and Clinical Medicine.85(2): 235-244.
Bailly C 2004. Active oxygen species and antioxidants in seed biology. Seed Science Research.14(2): 93-107.
Bailly C, Benamar A, Corbineau F & Côme D 1996. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiologia Plantarum.97(1): 104-110.
Barros J, Basch G & de Carvalho M 2005. Effect of reduced doses of a post-emergence graminicide mixture to control Lolium rigidum G. in winter wheat under direct drilling in Mediterranean environment. Crop Protection.24(10): 880-887.
Barroso A, Yamauti M & Alves P 2010. Interference between weed species and two bean cultivars in two times of sowing. Bragantia.69(3): 609-616.
Blackshaw R, Odonovan J, Harker K, Clayton G & Stougaard R 2006. Reduced herbicide doses in field crops: a review. Weed Biology and Management.6(1): 10-17.
Boström U & Håkan F 2002. Response of weeds and crop yield to herbicide dose decision-support guidelines. Weed Science 50, no. 2 50(2): 186-195.
Boulahia K, Carol P, Planchais S & Abrous-Belbachir O 2016. Phaseolus vulgaris L. seedlings exposed to prometryn herbicide contaminated soil trigger an oxidative stress response. Journal of Agricultural and Food Chemistry 64(16): 3150-3160.
Caverzan A, Casassola A & Brammer S 2016. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. SHANKER AK & SHANKER C. Abiotic and biotic stress in plants-Recent advances and future perspectives. In-Tech.
Czarnocka W & Karpiński S 2018. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radical Biology and Medicine 122: 4-20.
Délye C, Jasieniuk M & Le Corre V 2013. Deciphering the evolution of herbicide resistance in weeds. Trends in Genetics 29(11): 649-658.
Demirevska-Kepova K, Simova-Stoilova L, Stoyanova Z, Hölzer R & Feller U 2004. Biochemical changes in barley plants after excessive supply of copper and manganese. Environmental and Experimental Botany 52, no. 3 52(3): 253-266.
Dogan M 2005. Efficacies of reduced herbicide rates for weed control in maize (Zea mays L.) during critical period. Journal of Agronomy 4(1): 44-48.
Dubey R 2010. Metal toxicity, oxidative stress and antioxidative defense system in plants. Reactive Oxygen Species and Antioxidants in Higher Plants.15: 193-220.
Fanadzo M, Chiduza C & Mnkeni P 2010. Reduced dosages of atrazine and narrow rows can provide adequate weed control in smallholder irrigated maize (Zea mays L.) production in South Africa. African Journal of Biotechnology.9(45): 7660-7666.
Foyer C & Halliwell B 1976. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta.133(1): 21-25.
Foyer C & Noctor G 2005. Redox homeostasis and antioxidant signaling: a metabolic
interface between stress perception and physiological responses. The Plant Cell.17(7): 1866-1875.
Galeshi S, Torabi B, Resam G, RahemiKarizaki A & Barzegar A 2009. Stress management in plants. Publications Gorgan University of Agricultural Sciences and Natural Resources. 24-29.
Gill S & Tuteja N 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry.48(12): 909-930.
Gupta N, Gupta S & Kumar A 2001. Effect of water stress on physiological attributes and their relationship with growth and yield of wheat cultivars at different stages. Journal of Agronomy and Crop Science.186(1): 55-62.
Han Y & Wang C 2002. Physiological basis of bentazon tolerance in rice (Oryza sativa L.) lines. Weed Biology and Management.2(4): 186-193.
Hatzio K & Penner D 1985. Interactions of herbicides with other agrochemicals in higher plants. Reviews of weed science (USA). 5(2): 173-182.
Heap I 2014. Global perspective of herbicide‐resistant weeds. Pest Management Science.70(9): 1306-1315.
Hugie J, Bollero G, Tranel P & Riechers D 2008. Defining the rate requirements for synergism between mesotrione and atrazine in redroot pigweed (Amaranthus retroflexus). Weed Science.56(2): 265-270.
Jiang L & Yang. H 2009. Prometryne-induced oxidative stress and impact on antioxidant enzymes in wheat. Ecotoxicology and Environmental Safety.72(6): 1687-1693.
Jung S 2004. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Science.166(2): 459-466.
Kafi M, et al. 2009. Physiology of environmental stresses in plants, Mashhad: Academic Center for Education, Culture and Research. 87-96.
Knežević M, Đurkić M, Knežević I, Antonić O & Jelaska S 2003. Effects of tillage and reduced herbicide doses on weed biomass production in winter and spring cereals. Plant, Soil and Environment.49(9): 414-421.
Krausz R, Young B, Kapusta G & Matthews J 2001. Influence of weed competition and herbicides on glyphosate-resistant soybean (Glycine max). Weed Technology.15(3): 530-534.
Krivosheeva A, et al. 1996. Cold acclimation and photoinhibition of photosynthesis in Scots pine. Planta.200(3): 296-305.
Li H, et al. 2014. Overexpression of Pp14-3-3 from Pyrus pyrifolia fruit increases drought and salt tolerance in transgenic tobacco plant. Biologia.69(7): 880-887.
Merve A & Burcu S 2012. Changes in physiological parameters and some antioxidant enzymes activities of soybean (Glycine max L. Merr.) leaves under cadmium and salt stress. Journal of Stress Physiology & Biochemistry 8(4).
Mittler R 2002. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science.7(9): 405-410.
Mittler R 2017. Ros are good. Trends in Plant Science.22(1): 11-19.
Mühling K & Läuchli A 2003. Interaction of NaCl and Cd stress on compartmentation pattern of cations, antioxidant enzymes and proteins in leaves of two wheat genotypes differing in salt tolerance. Plant and Soil.253(1): 219-231.
Nakano Y & Kozi A 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant and Cell Physiology 22(5): 867-880.
Pan D, et al. 2017. Interactions between salicylic acid and antioxidant enzymes tilting the balance of H2O2 from photorespiration in non-target crops under halosulfuron-methyl stress. Pesticide Biochemistry and Physiology.143: 214-223.
Powles S & Yu Q 2010. Evolution in action: plants resistant to herbicides. Annual Review of Plant Biology.61: 317-347.
Rosales-Robles E, Sanchez-de-la-Cruz R, Salinas-Garcia J & Pecina-Quintero V 2005. Broadleaf weed management in grain sorghum with reduced rates of postemergence herbicides. Weed Technology 19(2): 385-390.
Tan Y, Zongsuo L, Hongbo S & Feng D 2006. Effect of water deficits on the activity of anti-oxidative enzymes and osmoregulation among three different genotypes of Radix Astragali at seeding stage. Colloids and Surfaces B: Biointerfaces.49(1): 60-65.
Triantaphylidès C & Michel H 2009. Singlet oxygen in plants: production, detoxification and signaling. Trends in Plant Science.14(4): 219-228.
Walker S, Medd R, Robinson G & Cullis B 2002. Improved management of Avena ludoviciana and Phalaris paradoxa with more densely sown wheat and less herbicide. Weed Research 42(4): 257-270.
Wang S, et al. 2004. Copper-induced stress and antioxidative responses in roots of Brassica juncea L. Botanical Bulletin of Academia Sinica.45.
Williams M & Nelson R 2014. Vegetable soybean tolerance to bentazon, fomesafen, imazamox, linuron, and sulfentrazone. Weed Technology.28(4): 601-607.
Xu J, Hengxia Y, Xiaojing L & Xia L 2010. Salt affects plant Cd-stress responses by modulating growth and Cd accumulation. Planta.231(2): 449-459.
Yordanova R, Christov K & Popova L 2004. Antioxidative enzymes in barley plants subjected to soil flooding. Environmental and Experimental Botany.51(2): 93-101.
Yuan J, Tranel P & Stewart Jr C 2007. Non-target-site herbicide resistance: a family business. Trends in plant science.12(1): 6-13.
Zhang J, Hamill A & Weaver S 1995. Antagonism and synergism between herbicides: trends from previous studies. Weed Technology. 86-90.
Zhang J, Lu Y, Zhang J, Tan L & Yang H 2014. Accumulation and toxicological response of atrazine in rice crops. Ecotoxicology and Environmental Safety.102: 105-112.
Zhang J, Weaver S & Hamill A 2000. Risks and reliability of using herbicides at below-labeled rates. Weed Technology.14(1): 106-115.
Zhang X, Ervin E, Evanylo G, Sherony C & Peot C 2005. Biosolids impact on tall fescue drought resistance. Journal of Residuals Science & Technology.2: 173-180.
Zhu J, Patzoldt W, Radwan O, Tranel P & Clough S 2009. Effects of photosystem-II-interfering herbicides atrazine and bentazon on the soybean transcriptome. Plant Genome.2(2): 191-205.
Type of article: orginal article |
Subject:
public specific
Received: 2019/12/2 | Published: 2020/07/27 | ePublished: 2020/07/27
Received: 2019/12/2 | Published: 2020/07/27 | ePublished: 2020/07/27
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
